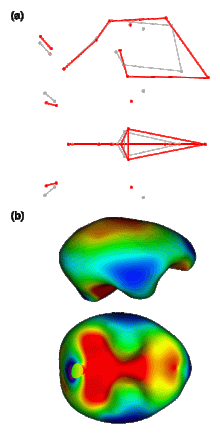
We have recently published a new paper regarding modularity and community detection in the context of human brain anatomical network analysis (Schuurman & Bruner, 2023a). Humans’ morphologically complex brain is spatially constrained by the physical interactions of its elements, and this aspect can be studied using anatomical network analysis. A crucial issue in this framework is modularity, assessed by means of community detection algorithms: the presence of groups of elements undergoing morphological changes in a concerted manner.
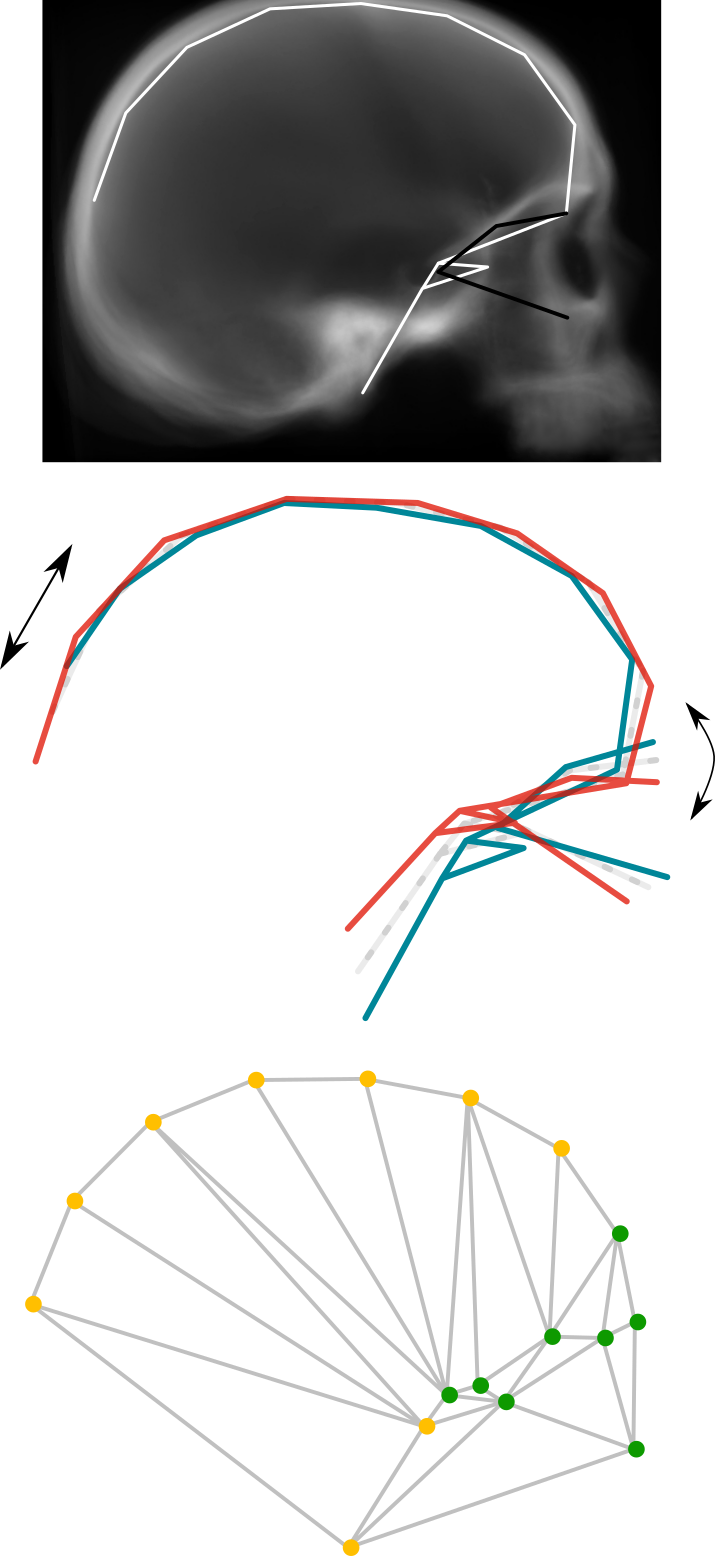
Previous works on the subject had produced mixed results: Bruner et al. (2019) and Bruner (2022) found longitudinal (anterior-posterior) modular partitions of the brain, while Schuurman & Bruner (2023b) found a vertical (superior-inferior) modular partition. In the present study, our aim was to test an array of community detection algorithms on a previously designed anatomical network model (Schuurman & Bruner, 2023b), offering a quantitative exploration of modularity to generate well-founded interpretations of the observed morphological organization of the human brain.
The algorithms that were examined comprise the combined method of the spin-glass model and simulated annealing algorithm (SG-SA), a partitional method with an automatic determination of the number of communities; the Louvain method, a heuristic method reliant on Q optimization with no a priori assumptions on community size or number; Infomap, an algorithm that focuses on information diffusion across the network; the generalized topological overlap measure (GTOM), a hierarchical clustering method based on the level of structural equivalence of the network’s nodes; and agglomerative hierarchical clustering based on maximal clique (EAGLE), which identifies initial subsets of completely connected nodes, then merges these subsets hierarchically according to their topological similarity and identifies the ideal partition using Q optimization.
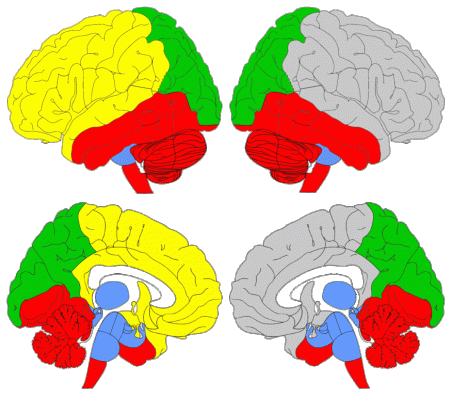
The algorithms that provide the highest quality partitions are SG-SA and the Louvain method. However, all five of them supply useful information. SG-SA, Louvain and Infomap reveal a clear vertical modular partition of the brain: they yield only two superior modules (these span regions as posterior as the middle occipital gyrus) and three inferior modules. Additionally, Informap goes as far as to subdivide the posterior inferior module vertically again, into a superior and an inferior posterior submodule. On the other hand, GTOM and EAGLE, show a more obvious longitudinal division: firstly, they separate the superior modules longitudinally (these only reach as far as the supramarginal gyrus), generating an additional, more posterior, superior submodule; and secondly, many regions incorporated in the inferior modules by SG-SA, the Louvain method and Infomap, are here considered to be part of the superior modules, contributing even further to this sense of anterior versus posterior. In other words, jointly, the community detection algorithms expose the simultaneous occurrence of a longitudinal and a vertical modular partition. This combination mirrors the morphological organization of the enveloping braincase, separated vertically by the distinct developmental processes of the cranial base and vault, and longitudinally by the particular morphogenetic environments of the three endocranial fossae. Overall, results suggest a level of concerted topological reciprocity in the spatial arrangement of craniocerebral anatomical components. Lastly, they posit questions concerning the degree to which structural constraints of the skull and the modular partition of the brain may channel both evolutionary and developmental trajectories.
Tim Schuurman